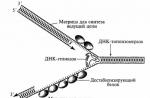
Virus reproduction
Viruses are characterized by a disjunctive (from disjuncus - disunited) method of reproduction-multiplication. Virus progeny results from the assembly of nucleic acids and protein subunits, which are synthesized separately by the host cell. The penetration of a virus into a cell and the reproduction of its own kind takes place in several phases: penetration into the host cell, synthesis of enzymes necessary for the replication of viral nucleic acids, synthesis of viral parts, assembly and composition of mature virions, exit of mature virions from the cell.
Phase I -- adsorption of the virion on the cell surface.
It proceeds in two stages: the first is nonspecific, when the virus is retained on the cell surface due to the appearance of opposite charges between individual sections of the cell membrane and the virus. This phase of the interaction of the virus with the cell is reversible, it is influenced by factors such as pH and salt composition of the medium.
The second stage is specific, when specific virus receptors and cell receptors that are complementary to each other interact. By chemical nature, cell receptors can be mucoproteins (or mucopolysaccharides) and lipoproteins. Different viruses are fixed on different receptors: influenza viruses, parainfluenza, adenoviruses - on mucoproteins, and tick-borne encephalitis, poliomyelitis viruses - on lipoproteins.
Phase II - the penetration of the virus into the cell. Electronoscopy observations of the process of penetration of viruses into cells sensitive to them showed that it is carried out through a mechanism resembling pinocytosis, or, as it is more commonly called, viropexis. At the site of adsorption of the virus, the cell wall is drawn into the cell, a vacuole is formed, in which the virion is found. In parallel, cellular enzymes (lipases and proteases) cause the deproteinization of the virion - the dissolution of the protein shell and the release of the nucleic acid.
Phase III - a latent period (the period of the eclipse - disappearance). During this period, it is impossible to determine the presence of an infectious virus in the cell either by chemical, or electron microscopic, or serological methods. Little is known about the nature of this phenomenon and its mechanisms. It is assumed that in the latent phase, the nucleic acid of the virus penetrates into the chromosomes of the cell and enters into complex genetic relationships with them.
Phase IV -- synthesis of virion components. In this phase, the virus and the cell are a single whole, the viral nucleic acid performs a genetic function, induces the formation of early proteins and changes the function of ribosomes. Early proteins are divided into:
a) inhibitory proteins (repressors) that suppress cell metabolism
b) enzyme proteins (polymerases) that provide the synthesis of viral nucleic acids.
The synthesis of nucleic acids and proteins proceeds non-simultaneously and in different structural parts of the cell. In viruses containing DNA or RNA, these processes have some differences and features.
Phase V is the formation of mature virions. The process of "assembly" of the virus is carried out as a result of the connection of the components of the virus particle. In complex viruses, cellular structures take part in this process and the inclusion of lipid, carbohydrate, and protein components of the host cell into the viral particle occurs.
The process of formation of virions begins after a certain time after the synthesis of their constituent components has begun. The duration of this period is quite variable and is predetermined by the nature of the virus - for RNA-containing viruses it is usually shorter than for DNA viruses. For example, the production of complete vaccinia virus particles begins approximately 5–6 hours after cell infection and continues for the next 7–8 hours, i.e., after the synthesis of viral DNA has already been completed.
Very strong bonds are formed between the nucleic acid and the corresponding protein subunits, as evidenced by the difficulty in separating the protein from the viral nucleic acid. The greater strength of the viral particle is given by its constituent carbohydrates and especially lipids.
The formation of virions, as well as the synthesis of virus components, occurs in different places in the cell, with the participation of various cellular structures. After the formation process is completed, a mature daughter virus particle is formed, which has all the properties of the parent virion. But sometimes the formation of so-called incomplete viruses is observed, which consist either only of a nucleic acid, or of a protein, or of viral particles, the formation of which has stopped at some intermediate stage.
Phase VI is the release of mature virions from the cell. There are two main mechanisms for the release of mature virions from the cell: 1) the release of the virion by budding. In this case, the outer envelope of the virion is derived from the cell membrane and contains both host cell material and viral material; 2) exit of mature virions from the cell through gaps in the membrane. These viruses do not have an outer shell. With such a mechanism for the release of viruses, the cell, as a rule, dies and a large number of virus particles appear in the medium.
Features of the reproduction of viruses
1. Periods of productive viral infection
2. Virus replication
3. Broadcast
1. A productive viral infection occurs in 3 periods:
The initial period includes the stages of adsorption of the virus on the cell, penetration into the cell, disintegration (deproteinization) or "undressing" of the virus. The viral nucleic acid was delivered to the appropriate cell structures and, under the action of lysosomal cell enzymes, is released from protective protein coats. As a result, a unique biological structure is formed: an infected cell contains 2 genomes (own and viral) and 1 synthetic apparatus (cellular);
After this, the second group of virus reproduction processes begins, including the middle and final periods, during which repression of the cellular and expression of the viral genome occur. Repression of the cellular genome is provided by low molecular weight regulatory proteins such as histones, which are synthesized in any cell. With a viral infection, this process is enhanced, now the cell is a structure in which the genetic apparatus is represented by the viral genome, and the synthetic apparatus is represented by the synthetic systems of the cell.
2. The further course of events in the cell is aimed at replication of the viral nucleic acid (synthesis of genetic material for new virions) and the implementation of the genetic information contained in it (synthesis of protein components for new virions). In DNA-containing viruses, both in prokaryotic and eukaryotic cells, viral DNA replication occurs with the participation of the cellular DNA-dependent DNA polymerase. In this case, in single-stranded DNA-containing viruses, a complementary strand is first formed - the so-called replicative form, which serves as a matrix for daughter DNA molecules.
3. The implementation of the genetic information of the virus contained in DNA occurs as follows: with the participation of DNA-dependent RNA polymerase, mRNAs are synthesized, which enter the ribosomes of the cell, where virus-specific proteins are synthesized. In double-stranded DNA-containing viruses, the genome of which is transcribed in the cytoplasm of the host cell, this is its own genomic protein. Viruses whose genomes are transcribed in the cell nucleus use the cellular DNA-dependent RNA polymerase contained there.
In RNA-containing viruses, the processes of replication of their genome, transcription and translation of genetic information are carried out in other ways. Replication of viral RNA, both minus and plus strands, is carried out through the replicative form of RNA (complementary to the original), the synthesis of which is provided by RNA-dependent RNA polymerase - this is a genomic protein that all RNA-containing viruses have. The replicative form of RNA of minus-strand viruses (plus-strand) serves not only as a template for the synthesis of daughter viral RNA molecules (minus-strands), but also performs the functions of mRNA, i.e. goes to ribosomes and ensures the synthesis of viral proteins (translation ).
In plus-strand RNA-containing viruses, the function of translation is performed by its copies, the synthesis of which is carried out through the replicative form (minus-strand) with the participation of viral RNA-dependent RNA polymerases.
Some RNA viruses (reoviruses) have a completely unique transcription mechanism. It is provided by a specific viral enzyme - reverse transcriptase (reverse transcriptase) and is called reverse transcription. Its essence lies in the fact that at first a transcript is formed on the viral RNA matrix with the participation of reverse transcription, which is a single strand of DNA. On it, with the help of cellular DNA-dependent DNA polymerase, the second strand is synthesized and a double-stranded DNA transcript is formed. From it, in the usual way, through the formation of i-RNA, the information of the viral genome is realized.
The result of the described processes of replication, transcription and translation is the formation of daughter viral nucleic acid molecules and viral proteins encoded in the genome of the virus.
This is followed by the third, final period of interaction between the virus and the cell. New virions are assembled from the structural components (nucleic acids and proteins) on the membranes of the cytoplasmic reticulum of the cell. A cell whose genome has been repressed (suppressed) usually dies. The newly formed virions passively (as a result of cell death) or actively (by budding) leave the cell and find themselves in its environment.
Thus, the synthesis of viral nucleic acids and proteins and the assembly of new virions occur in a certain sequence (separated in time) and in different cell structures (separated in space), and therefore the method of reproduction of viruses was called disjunctive (separated). With an abortive viral infection, the process of interaction of the virus with the cell is interrupted for one reason or another before the suppression of the cellular genome has occurred. Obviously, in this case, the genetic information of the virus will not be realized and the reproduction of the virus does not occur, and the cell retains its functions unchanged. virion cell virus
During a latent viral infection, both genomes function simultaneously in the cell, while during virus-induced transformations, the viral genome becomes part of the cellular one, functions and is inherited along with it.
Bibliography
1. V. A. Sergeev et al., Veterinary Virology. - Moscow, 2002.
2. Virology. Edited by Fields B., Knight D., vols. 1-3, M., 1989.
3. Gosmanov R.G., Kolychev N.M. Veterinary virology. M.: Kolos. - 2003.
4. Belousova R.V., Preobrazhenskaya E.A., Tretyakova I.V. Veterinary virology: A textbook for universities (ed. Belousova R.V.). - M.: Kolos. - 2007
Synthesis of a group of early proteins ( repressors of cellular metabolism, virus-specific polymerases), Synthesis of virus-specific proteins and NA, Synthesis of structural (late) proteins of the virus, Formation of mature virions This synthesis is based on the same mechanism as in protein synthesis in normal cells. In RNA-containing picornaviruses, the mRNA function is performed by plus strands. Their single-stranded virion RNA is translated to form a single giant polypeptide, which is then cleaved into individual functional proteins. In the synthesis of a complete viral protein, the translation of all viral RNA genes is expressed in time. In ortho-, rhabdo- and paramyxes, the virion single-stranded RNA is not translated, but the plus-strand complementary to it is translated, therefore the synthesis of viral specific proteins begins after the formation of viral mRNA (plus-strand), which is complementary to the virion RNA-Minus-strands are synthesized on the plus-strand. strands of virion RNA-dependent polymerase (RNA transcriptase), which is part of the virion as a structural component. The mRNAs synthesized by the virion polymerase are monocistronic and much shorter than the virion RNA. During a viral infection, cell polysomes disintegrate and virus-specific polysomes form. The synthesis of virus-specific protein depends on the synthesis of viral mRNA, but also affects it: if protein synthesis is disturbed, the newly formed mRNA is overstocked at the sites of its synthesis and its further synthesis is inhibited. Viral proteins during infection are synthesized in excess quantities than are required for the formation of an infectious virus. For example, in cells infected with herpes viruses, only about 35% of the total mass of virus-specific proteins synthesized in cells is included in the viral progeny. In most viruses, protein synthesis is carried out in the cytoplasm; there is doubt about the nuclear localization of protein synthesis in some viruses. It is known that viral proteins can be synthesized in some structures and accumulated in others. The mechanisms responsible for the migration of viral proteins into the nucleus have not been elucidated. It is only known that the absence of arginine in the medium leads to suppression of the migration of structural proteins of the herpes virus from the site of their synthesis (cytoplasm) to the site of assembly of virions (nucleus), although the synthesis of DNA and protein of the virus is not impaired. At different stages of the infectious cycle, one or another group of virus-specific proteins can be predominantly formed. Their speed is regulated either at the level of transcription (with the formation of mRNA) or at the level of translation (reading of mRNA on ribosomes). In an infected cell, mRNAs from different viral genes disproportionately accumulate. The mechanism of this disproportionality lies in the virus particle itself. The same mechanism determines the different efficiency of formation of different proteins. A standard viral particle contains one RNA molecule and up to 10,000 protein molecules. In addition to structural proteins, non-structural (but encoded by viral RNA) proteins can also be synthesized in an infected cell. Along with the synthesis of proteins in the cell during the reproduction of the influenza virus, the synthesis of carbohydrate chains that make up glycoproteins also occurs. Attachment of carbohydrates is carried out with the help of transferases, which are cellular enzymes. Lipid synthesis is also carried out by the cell. The viral envelope is formed by the incorporation of lipids from the plasma membrane of the host cell. The synthesis of viral nucleic acids and virus-specific proteins occurs almost simultaneously and is at least 1 hour ahead of the maturation of viral particles.
Stages of virus-cell interaction: adsorption and penetration of bacteriophages.
Adsorption: non-specific, specific.
Penetration: Injection (shells remain - “shadows” of phages), Transfection.
+ Question #6
Stages of interaction between a virus and a cell: penetration and spread of phytopathogenic viruses.
Adsorption: No. Penetration: Contact (through injury) From mother plant to daughter Arthropod vectors. Reproduction: Synthesis of a group of early proteins ( repressors of cellular metabolism, virus-specific polymerases) Synthesis of virus-specific proteins and NA. Synthesis of structural (late) proteins of the virus. Formation of mature virions Assembly: automatic. Exit:
Stages of interaction of the virus with the cell: types of release of viral particles from the cell.
Animal viruses: Extrusion of sections of the cytoplasm. Individual output virions or their groups. Plant viruses: through intercellular connections. The cells produce the virus without being lysed. Bacteriophages: After cell death (lytic type).
Types of interaction between the virus and the cell: productive type.
Productive infection- the formation of new viral particles.
lytic– the infected cell can die, thus forming a large amount of the virus. Bacteriophages that form a new generation of phage particles in infected cells, which leads to lysis (destruction) of the bacterial cell, are called virulent phages. Some bacteriophages do not replicate inside the host cell. Instead, their nucleic acid is incorporated into the host's DNA, forming with it a single molecule capable of replication. Such phages are called temperate phages, or prophages. The prophage has no lytic effect on the host cell and replicates together with the cellular DNA during division. Bacteria containing prophage are called lysogenic. They show resistance to the phage contained in them, as well as to other phages close to it. The connection between a prophage and a bacterium is very strong, but it can be broken under the influence of inducing factors (UV - rays, ionizing radiation, chemical mutagens). persistent- the cell continues to live and divide, synthesizing small amounts of the virus. characteristic of animal viruses. Forms of persistent infection: latent(herpes), chronic(hepatitis), slow infection(HIV).
Types of interaction between the virus and the cell: lysogenic type.
Integrative infection- the genome of the virus is integrated into the genome of the host cell and replicates along with it. There is no virus production. Virus carrying. temperate phages - able to lysogenize infected bacteria. Virulent phages - cannot lysogenize bacteria. Prophage - phage that is transmitted to daughter cells during division.
Classification of viruses.
Viruses are divided into 6 classes according to the structure of the nucleic acid: Double stranded DNA, Single stranded DNA, Double stranded RNA, Single stranded RNA "+" strand, Single stranded RNA "-" strand, Retroviruses The Baltimore classification of viruses is based on the mechanism of mRNA formation. Viruses must synthesize mRNA from their own genomes in order to form proteins and replicate their nucleic acid, but each family of viruses has its own mechanism for doing this. Viral genomes can be single-stranded (ss) or double-stranded (ds), contain DNA or RNA, and may or may not use reverse transcriptase. In addition, single-stranded RNA viruses can have positive (+) or negative (-) strand RNA as part of their genome.
This system includes seven main groups:(I) Viruses containing double-stranded DNA and lacking an RNA stage (eg, herpesviruses, poxviruses, papovaviruses, mimiviruses). (II) Viruses containing a single-stranded DNA molecule (for example, parvoviruses). In this case, the DNA is always positive polarity. (III) Viruses containing double-stranded RNA (eg rotaviruses). (IV) Viruses containing a positive polarity single-stranded RNA molecule (eg picornaviruses, flaviviruses). (V) Viruses containing a single-stranded RNA molecule of negative or dual polarity (eg, orthomyxoviruses, filoviruses). (VI) Viruses containing a single-stranded positive RNA molecule and having in their life cycle the stage of DNA synthesis on an RNA template, retroviruses (for example, HIV). (VII) Viruses containing double-stranded DNA and having in their life cycle the stage of DNA synthesis on an RNA template, retroid viruses (for example, hepatitis B virus). Further division is made on the basis of such features as the structure of the genome (the presence of segments, a circular or linear molecule), genetic similarity with other viruses, the presence of a lipid membrane, the taxonomic affiliation of the host organism, and so on.
Table of contents of the subject "Virology. Reproduction of viruses. Genetics of viruses.":1. Virology. History of virology. Chamberlain. RU. Pasteur. Ivanovsky.
2. Reproduction of viruses. Reproduction of +RNA viruses. Picornaviruses. reproduction of picornaviruses.
3. Togaviruses. Reproduction of togaviruses. Retroviruses. reproduction of retroviruses.
5. Reproduction of DNA viruses. Replication cycle of DNA-containing viruses. Reproduction of papovaviruses. Reproduction of adenoviruses.
6. Reproduction of herpes viruses. The replication cycle of herpesviruses. Poxviruses. Reproduction of poxviruses.
7. Reproduction of the hepatitis B virus. Replication cycle of the hepatitis B virus.
8. Genetics of viruses. Characteristics of viral populations. Gene pool of viral populations.
9. Mutations of viruses. Spontaneous mutations of viruses. Induced mutations of viruses. The manifestation of virus mutations in the phenotype.
10. Genetic interactions between viruses. Recombination and redistribution of genes by viruses. Exchange of genome fragments by viruses. Antigenic shift.
-RNA viruses enter the cell by fusion (paramyxoviruses) or viropexis (rhabdo- and orthomyxoviruses). For effective reproduction, viral RNA must be converted into +RNA, an analogue of cellular mRNA (Fig. 5-3).
Rice. 5-3. Reproductive cycle -RNA-containing viruses. The penetration of the virus into the cell occurs after its adsorption and fusion with the cell membrane (1). After the release of the viral -RNA, +RNA is synthesized on the -RNA template, catalyzed by the RNA-dependent RNA-pol imerase, which is part of the virion (2), which leads to the formation of full and short filaments. Short +RNA strands are involved in the synthesis of enzymes and proteins for daughter populations (3). Among the latter, the M protein (4) and envelope glycoproteins, which are incorporated into the cell wall at the stages preceding budding, are of particular importance. The complete +RNA chain serves as a template for the synthesis of -RNA molecules of daughter populations (5). The virions of the daughter populations are collected on sections of the cell membrane modified with the M protein (6) and released by budding, capturing its fragment, which later serves as a supercapsid (7).-RNA viruses. Reproduction of -RNA viruses
early stage of reproduction. After the release of the genome, viral transcriptase (RNA dependent RNA polymerase) triggers +RNA synthesis. In this case, the viral ribonucleoprotein (that is, RNA and internal proteins) serves as a “template” for viral transcriptase. As a result, complete and short molecules-copies of +RNA are formed.
late reproductive stage. Full plus strands serve as templates for the synthesis of RNA molecules that make up the genomes of the daughter population. Short plus filaments are involved in the synthesis of enzymes and proteins. Viral proteins (hemagglutinin and neuraminidase) interact with sections of the cell membrane. Viral M-proteins (matrix proteins) are also adsorbed there. They exhibit pronounced hydrophobicity due to the content of up to 75% of neutral amino acids. This property gives them the ability to interact with proteins and lipids of cell membranes and to mediate the assembly of viral particles. On the one hand, the M-protein recognizes the sites of inclusion of viral glycoproteins in the membrane, on the other hand, it is specifically recognized by the nucleocapsid and binds to it. The assembly of daughter populations is completed after the attachment of the nucleocapsid to the cell membrane. Their release occurs by budding through the modified sections of the membrane. Budding viral particles capture its fragments, which later serve as supercapsids.

Reproduction of viruses with double-stranded RNA
Double-stranded RNA viruses are represented by the Reoviridae family (reo- and rotaviruses). They do not have a supercapsid and are organized according to the type of cubic symmetry. An RNA-dependent RNA polymerase is associated with viral RNA. Viruses are distinguished by an extended reproductive cycle and a tendency to accumulate products of virus-specific synthesis inside cells. After the release of the genome in the cytoplasm of cells, RNA polymerase synthesizes mRNA molecules (+RNA on one strand -RNA. As a result, up to 11 functional mRNA molecules are formed, corresponding in size to 11 segments of one strand -RNA. The molecules are translated into 11 primary polypeptide products. Their subsequent cleavage leads to the formation of up to 16 secondary polypeptides in infected cells.Seven primary and two secondary polypeptides are part of the viral particles, the remaining primary and secondary polypeptides perform catalytic and regulatory functions.In parallel, the viral RNA polymerase synthesized during translation triggers the synthesis of negative strands on the matrix +RNA, followed by their connection into a double-stranded RNA molecule.The release of the resulting virions is accompanied by cell death.
The reproduction of the virus in the cell occurs in several phases (Fig. 7):
The first phase is the adsorption of the virus on the cell surface that is sensitive to this virus.
The second phase is the penetration of the virus into the host cell by viropexis.
The third phase is the "undressing" of virions, the release of the nucleic acid of the virus from the supercapsid and capsid. In a number of viruses, the penetration of the nucleic acid into the cell occurs by the fusion of the virion envelope and the host cell. In this case, the second and third phases are combined into one.
Depending on the type of nucleic acid, this process occurs as follows.
1. Reproduction occurs in the nucleus: adenoviruses, herpes, papo-vaviruses. Use DNA-dependent RNA - cell polymerase.
2. Reproduction occurs in the cytoplasm: viruses have their own DNA-dependent RNA polymerase. RNA-containing.
1. Riboviruses with a positive genome (plus-nitium): picor-
na-, toga-, coronaviruses. There is no transcription.
RNA -> protein
2. Genome-negative riboviruses (minus-threaded): influenza,
measles, mumps, ortho-, paramyxoviruses.
(-)RNA - > mRNA - > protein (mRNA complementary (-) RNA) This process takes place with the participation of a special viral enzyme - virion RNA-dependent RNA polymerase (there cannot be such an enzyme in a cell).
3. Retroviruses
(-) RNA -> DNA -> mRNA -> protein (and RNA is homologous to RNA) In this case, the process of forming DNA based on (-) RNA is possible with the participation of the enzyme - RNA-dependent DNA polymerase (reverse transcriptase or reverse transcriptase)
The fourth phase is the synthesis of virion components. The nucleic acid of the virus is produced by replication. The information of the viral mRNA is translated to the ribosomes of the cell, and a virus-specific protein is synthesized in them.
The fifth phase is the assembly of the virion. Nucleocapsids are formed by self-assembly.
The sixth phase is the release of virions from the cell. Simple viruses, such as the polio virus, destroy the cell when they exit. Complex viruses, such as the influenza virus, exit the cell by budding. The outer shell of the virus (supercapsid) is formed during the release of the virus from the cell. The cell during this process remains alive for some time.
The described types of interaction of the virus with the cell are called productive, as they lead to the production of mature virions.
Another way - integrative - is that after the penetration of the virus into the cell and "undressing" the viral nucleic acid integrates into the cell genome, that is, it is integrated into the cell chromosome in a certain place and then replicated along with it in the form of the so-called provirus. For DNA- and RNA-containing viruses, this process takes place in different ways. In the first case, viral DNA integrates into the cellular genome. In the case of RNA-containing viruses, reverse transcription occurs first: DNA is formed on the viral RNA template with the participation of the "reverse transcriptase" enzyme, which is integrated into the cellular genome. The provirus carries additional genetic information, so the cell acquires new properties. Viruses capable of this type of interaction with a cell are called integrative. Integrative viruses include some oncogenic viruses, hepatitis B virus, herpes virus, human immunodeficiency virus, temperate bacteriophages.
The reproduction of the virus in the cell occurs in several phases:
The first phase is the adsorption of the virus on the cell surface that is sensitive to this virus.
The second phase is the penetration of the virus into the host cell by viropexis.
The third phase is the "undressing" of virions, the release of the nucleic acid of the virus from the supercapsid and capsid. In a number of viruses, the penetration of the nucleic acid into the cell occurs by the fusion of the virion envelope and the host cell. In this case, the second and third phases are combined into one.
Depending on the type of nucleic acid, this process occurs as follows.
Reproduction occurs in the nucleus: adenoviruses, herpes, papovaviruses. Use DNA-dependent RNA - cell polymerase.
Reproduction occurs in the cytoplasm: viruses have their own DNA-dependent RNA polymerase.
Riboviruses with a positive genome (plus-nitium): picorna-, toga-, coronaviruses. There is no transcription.
RNA -> protein
Riboviruses with a negative genome (minus-strand): influenza, measles, parotitis, ortho-, paramyxoviruses.
(-)RNA -> mRNA -> protein (mRNA is complementary to (-)RNA). This process takes place with the participation of a special viral enzyme - virion RNA-dependent RNA polymerase (there cannot be such an enzyme in a cell).
Retroviruses
(-)RNA -> DNA -> mRNA -> protein (and RNA is homologous to RNA). In this case, the process of DNA formation on the basis of (-) RNA is possible with the participation of the enzyme - RNA-dependent DNA polymerase (reverse transcriptase or reverse transcriptase)
The fourth phase is the synthesis of virion components. The nucleic acid of the virus is produced by replication. The information of the viral mRNA is translated to the ribosomes of the cell, and a virus-specific protein is synthesized in them.
The fifth phase is the assembly of the virion. Nucleocapsids are formed by self-assembly.
The sixth phase is the release of virions from the cell. Simple viruses, such as the polio virus, destroy the cell when they exit. Complex viruses, such as the influenza virus, exit the cell by budding. The outer shell of the virus (supercapsid) is formed during the release of the virus from the cell. The cell during this process remains alive for some time.
The described types of interaction of the virus with the cell are called productive, as they lead to the production of mature virions.
Another way - integrative - is that after the penetration of the virus into the cell and "undressing" the viral nucleic acid integrates into the cell genome, that is, it is integrated into the cell chromosome in a certain place and then replicated along with it in the form of the so-called provirus. For DNA- and RNA-containing viruses, this process takes place in different ways. In the first case, viral DNA integrates into the cellular genome. In the case of RNA-containing viruses, reverse transcription occurs first: DNA is formed on the viral RNA template with the participation of the "reverse transcriptase" enzyme, which is integrated into the cellular genome. The provirus carries additional genetic information, so the cell acquires new properties. Viruses capable of this type of interaction with a cell are called integrative. Integrative viruses include some oncogenic viruses, hepatitis B virus, herpes virus, human immunodeficiency virus, temperate bacteriophages.
In addition to ordinary viruses, there are prions - protein infectious particles that do not contain nucleic acid. They look like fibrils, up to 200 nm in size. They cause slow infections in humans and animals with brain damage: Creutzfeldt-Jakob disease, kuru, scrapie and others.
Methods for the indication of viruses in the test material.
The reproduction of viruses in cell cultures is judged by their cytopathic action (CPE), which has a different character depending on the type of virus, by plaque formation on a cell monolayer covered with a thin agar layer, erythrocyte hemadsorption, and other tests.
Thus, the indication of viruses is performed microscopically by the presence of CPP, plaque formation on the cell monolayer, hemadsorption of erythrocytes added to the cell culture of the virus, as well as in the hemagglutination reaction with the tested virus-containing material. The hemagglutination reaction is caused by viruses containing hemagglutinin in their capsid or supercapsid.